Tstk voolab sisse. Trikarboksüülhappe tsükkel (TCA tsükkel)
- Üldine idee. Tsükli tsükli etappide tunnused.
- TFC lõpptooted.
- TCA tsükli bioloogiline roll.
- TCA tsükli reguleerimine.
- Häired keskküttesüsteemi töös.
· ÜLDVAADE. CTC STAADIUMIDE OMADUSED
Trikarboksüülhappe tsükkel (TCA tsükkel) on peamine, tsükliline, metaboolne rada, milles toimub aktiivse äädikhappe ja mõnede teiste süsivesikute, lipiidide, valkude lagunemisel tekkivate ühendite oksüdatsioon ning mis varustab hingamisahelat vähendatud koensüümidega.
CTK avati 1937. aastal G. Krebs. Ta võttis kokku sel ajal kättesaadavad eksperimentaalsed uuringud ja koostas protsessi täieliku diagrammi.
TCA tsükli reaktsioonid jätkuvad mitokondrites aeroobsetes tingimustes.
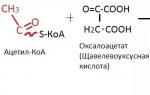
Tsükli alguses (joonis 6) kondenseerub aktiivne äädikhape (atsetüül-CoA) oksaloäädikhappega (oksaloatsetaat), moodustades sidrunhape (tsitraat). Seda reaktsiooni katalüüsitakse tsitraadi süntaas .
Seejärel tsitraat isomeriseeritakse isotsitraat. Tsitraadi isomerisatsioon viiakse läbi dehüdratsiooniga, moodustades cis-akonitaati ja selle järgneva hüdratatsiooni. Mõlema reaktsiooni katalüüs annab akonitaas .
Tsükli 4. etapis toimub isotsitraadi oksüdatiivne dekarboksüülimine tsükli mõjul. isotsitraatdehüdrogenaas (ICDG) haridusega a-ketoglutaarhape, NADH(H+) või NADPH(H+) ja CO2 . NAD-sõltuv IDH paikneb mitokondrites ja NADP-sõltuv ensüüm esineb mitokondrites ja tsütoplasmas.
5. etapis toimub a-ketoglutaraadi oksüdatiivne dekarboksüülimine koos moodustumisega. aktiivne merevaikhape (suktsinüül-CoA), NADH(H) ja CO2. Seda protsessi katalüüsitakse a-ketoglutaraadi dehüdrogenaasi kompleks , mis koosneb kolmest ensüümist ja viiest koensüümist. Ensüümid: 1) a-ketoglutaraatdehüdrogenaas, mis on seotud koensüümiga TPP; 2) transsuktsinülaas, mille koensüümiks on lipoehape;
3) FAD-ga seotud dihüdrolipoüüldehüdrogenaas. A-ketoglutaraadi dehüdrogenaaside töös
See kompleks hõlmab ka koensüüme CoA-SH ja NAD.
6. etapis lõhustatakse suktsinüül-CoA kõrge energiasisaldusega tioesterside koos SKT fosforüülimisega. Moodustuvad merevaikhape (suktsinaat) Ja GTP (substraadi fosforüülimise tasemel). Reaktsioon katalüüsitakse suktsinüül-CoA süntetaas (suktsinüültiokinaas) . GTP fosforüülrühma saab üle kanda ADP-le: GTP + ADP ® SKT + ATP. Reaktsiooni katalüüsitakse ensüümi nukleosiiddifosfokinaasi osalusel.
7. etapil suktsinaat oksüdeerub toimel suktsinaatdehüdrogenaas haridusega fumaraatja FADN 2.
8. etapil fumaraadi hüdrataas tagab vee lisamise moodustunud fumaarhappele L-õunhape (L-malaat).
L-malaat 9. staadiumis mõju all malaadi dehüdrogenaas oksüdeerub kuni oksaloatsetaat, tekib ka reaktsioon NADH(H+). Metaboolne rada sulgub oksaloatsetaadil ja uuesti kordab ennast, ostmine tsükliline iseloomu.
Riis. 6. Trikarboksüülhappe tsükli reaktsioonide skeem.
· LÕPPTOOTED
Üldine CTC võrrand on järgmisel kujul:
// KOHTA
CH 3 – C~ S-CoA + 3 NAD + + FAD + ADP + H 3 PO 4 + 3 H 2 O ®
® 2 CO 2 + 3 NADH(H +) + FADH 2 + ATP + CoA-SH
Seega on tsükli lõppsaadused (1 käibe kohta) redutseeritud koensüümid - 3 NADH (H +) ja 1 FADH 2, 2 molekuli süsinikdioksiidi, 1 molekul ATP ja 1 molekul CoA. - SH.
· TCA tsükli BIOLOOGILINE ROLL
Esineb Krebsi tsükkel integratsioon, amfiboolne (st kataboolne ja anaboolne), energia ja vesiniku doonori roll.
Integratsioon roll on selles, et TTC on lõplik ühine oksüdatsioonirada kütusemolekulid – süsivesikud, rasvhapped ja aminohapped.
Toimub TsTK-s atsetüül-CoA oksüdatsioon onkataboolnerolli.
Anaboolsed tsükli roll on see, et see varustab vahetooted Sest biosünteetiline protsessid. Näiteks kasutatakse sünteesimiseks oksaloatsetaati aspartaat, a-ketoglutaraat – hariduseks glutamaat, suktsinüül-CoA – sünteesiks heem.
Üks molekul ATP moodustatakse TCA-s tasemel substraadi fosforüülimine on energiat rolli.
Vesiniku doonor Roll on selles, et TCA tsükkel tagab koensüümide vähenemise NADH(H+) ja FADH 2 hingamisahel, milles toimub vesiniku oksüdatsioon nendest koensüümidest veeks koos ATP sünteesiga. Kui TCA tsüklis oksüdeeritakse üks atsetüül-CoA molekul, moodustub 3 NADH(H +) ja 1 FADH 2
ATP saagis atsetüül-CoA oksüdeerimisel on 12 ATP molekuli (1 ATP TCA tsüklis substraadi fosforüülimise tasemel ja 11 ATP molekuli 3 molekuli NADH(H +) ja 1 molekuli FADH 2 oksüdatsiooni ajal hingamisahel oksüdatiivse fosforüülimise tasemel).
· TCA TÜKLI REGULEERIMINE
Keskküttesüsteemi töökiirus on täpselt reguleeritud vajadustele rakud ATP-s, st. Krebsi tsükkel on seotud hingamisahelaga, mis toimib ainult aeroobsetes tingimustes. Tsükli oluline reguleeriv reaktsioon on tsitraadi süntees atsetüül-CoA-st ja oksaloatsetaadist, mis toimub tsitraadi süntaas. Kõrge ATP tase pärsib see ensüüm. Tsükli teine regulatiivne reaktsioon on isotsitraatdehüdrogenaas. ADP ja NAD + aktiveerida ensüüm, NADH(H+) ja ATP pärssida. Kolmas regulatiivne reaktsioon on a-ketoglutaraadi oksüdatiivne dekarboksüülimine. NADH(H+), suktsinüül-CoA ja ATP inhibeerivad a-ketoglutaraadi dehüdrogenaas.
· CTK TÖÖ HÄIRED
Rikkumine Tsentraalse tsirkulatsioonisüsteemi toimimine võib olla seotud:
atsetüül-CoA puudumisega;
Oksalatsetaadi puudumisega (see moodustub püruvaadi karboksüülimisel ja viimane omakorda süsivesikute lagunemisel). Süsivesikute toitumise tasakaalustamatus toob kaasa atsetüül-CoA kaasamise ketogeneesi (ketokehade moodustumine), mis põhjustab ketoosi;
Ensüümide aktiivsuse rikkumisega, mis on tingitud vitamiinide puudumisest, mis on osa vastavatest koensüümidest (vitamiini B 1 puudumine põhjustab TPP puudust ja a-ketoglutaraadi dehüdrogenaasi kompleksi talitlushäireid; vitamiin B 2 põhjustab FAD puudulikkust ja suktsinaatdehüdrogenaasi aktiivsuse rikkumist; vitamiini B 3 puudumine põhjustab koensüümi atsüülimise CoA-SH puudulikkust ja a-ketoglutaraatdehüdrogenaasi kompleksi aktiivsuse halvenemist; vitamiin B5 põhjustab NAD puudumist ja isotsitraatdehüdrogenaasi, a-ketoglutaraatdehüdrogenaasi kompleksi ja malaatdehüdrogenaasi aktiivsuse halvenemist; lipoehappe puudumine põhjustab ka a-ketoglutaraadi dehüdrogenaasi kompleksi talitlushäireid);
Hapnikupuuduse korral (hemoglobiini süntees ja hingamisahela toimimine on häiritud ning akumuleeruv NADH (H +) toimib sel juhul isotsitraatdehüdrogenaasi ja a-ketoglutaraadi dehüdrogenaasi kompleksi allosteerilise inhibiitorina)
· Kontrollküsimused
Rääkisin sellest, mis see tegelikult on, miks Krebsi tsüklit vaja on ja mis koha see ainevahetuses hõivab. Läheme nüüd selle tsükli reaktsioonide juurde.
Teen kohe broneeringu – minu jaoks isiklikult oli reaktsioonide päheõppimine täiesti mõttetu tegevus, kuni ma ülaltoodud küsimused ära sorteerisin. Aga kui olete teooriast juba aru saanud, soovitan liikuda edasi praktika juurde.
Krebsi tsükli kirjutamiseks näete palju võimalusi. Kõige tavalisemad valikud on midagi sellist:
Kuid mulle tundus kõige mugavam viis, kuidas kirjutada reaktsioone autorite T. T. Berezovi vanast heast biokeemia õpikust. ja Korovkina B.V.
Esimene reaktsioon
Juba tuttavad atsetüül-CoA ja oksaloatsetaat ühinevad ja muutuvad tsitraadiks, st sidrunhape.

Teine reaktsioon
Nüüd võtame sidrunhappe ja keerame seda isotsitrhape. Selle aine teine nimi on isotsitraat.
Tegelikult on see reaktsioon mõnevõrra keerulisem, läbi vahepealse etapi - cis-akoniithappe moodustumise. Kuid otsustasin seda lihtsustada, et see teile paremini meelde jääks. Vajadusel saate siia lisada puuduva sammu, kui kõik muu meelde tuleb.
Sisuliselt vahetasid kaks funktsionaalrühma lihtsalt kohad.

Kolmas reaktsioon
Niisiis, meil on isotsitrhape. Nüüd tuleb see dekarboksüülida (see tähendab, et COOH eemaldatakse) ja dehüdrogeenida (see tähendab, et H eemaldatakse). Saadud aine on a-ketoglutaraat.
See reaktsioon on märkimisväärne HADH 2 kompleksi moodustumisel. See tähendab, et NAD transporter kogub hingamisahela käivitamiseks vesinikku.
Mulle meeldib Berezovi ja Korovkini õpikus toodud versioon Krebsi tsükli reaktsioonidest just seetõttu, et reaktsioonides osalevad aatomid ja funktsionaalrühmad on kohe selgelt nähtavad.

Neljas reaktsioon
Jällegi, nikotiini amiid-adeniini dinukleotiid töötab nagu kellavärk, see tähendab ÜLAL. See kena kandja tuleb siia, nagu ka viimases etapis, haarama vesinikku ja kandma seda hingamisahelasse.
Muide, saadud aine on suktsinüül-CoA, ei tohiks teid hirmutada. Suktsinaat on merevaikhappe teine nimetus, mis on teile tuttav bioorgaanilise keemia päevilt. Succinyl-Coa on merevaikhappe ja koensüüm-A ühend. Võime öelda, et see on merevaikhappe ester.

Viies reaktsioon
Eelmises etapis ütlesime, et suktsinüül-CoA on merevaikhappe ester. Ja nüüd saame sama merevaikhape st suktsinaat, suktsinüül-CoA-st. Äärmiselt oluline punkt: see on selles reaktsioonis substraadi fosforüülimine.
Fosforüülimine üldiselt (see võib olla oksüdatiivne ja substraat) on fosforirühma PO 3 lisamine GDP-le või ATP-le, et saada täielikku GTF või vastavalt ATP. Substraat erineb selle poolest, et see sama fosforirühm rebitakse ära igast seda sisaldavast ainest. Lihtsamalt öeldes kantakse see SUBSTRATE'ilt HDF-i või ADP-sse. Sellepärast nimetatakse seda "substraadi fosforüülimiseks".
Veel kord: substraadi fosforüülimise alguses on meil difosfaadi molekul - guanosiindifosfaat või adenosiindifosfaat. Fosforüülimine seisneb selles, et kahe fosforhappejäägiga molekul – HDP või ADP – „lõpetatakse” kolme fosforhappejäägiga molekuliks, et saada guanosiin-trifosfaat või adenosiin-TRIfosfaat. See protsess toimub suktsinüül-CoA muundamisel suktsinaadiks (st merevaikhappeks).
Diagrammil näete tähti F (n). See tähendab "anorgaanilist fosfaati". Anorgaaniline fosfaat kantakse substraadist HDP-sse, nii et reaktsioonisaadused sisaldavad head, täielikku GTP-d. Vaatame nüüd reaktsiooni ennast:

Kuues reaktsioon
Järgmine transformatsioon. Seekord muutub merevaikhape, mille saime viimases etapis fumaraat, pange tähele uut kaksiksidet.
Diagramm näitab selgelt, kuidas see reaktsioonis osaleb FAD: See väsimatu prootonite ja elektronide kandja kogub vesinikku ja tõmbab selle otse hingamisahelasse.

Seitsmes reaktsioon
Oleme juba lõpusirgel. Krebsi tsükli eelviimane etapp on reaktsioon, mis muudab fumaraadi L-malaadiks. L-malaat on teine nimi L-õunhape, tuttav bioorgaanilise keemia kursuselt.
Kui vaatate reaktsiooni ennast, näete, et esiteks on see mõlemas suunas ja teiseks on selle olemus hüdratsioon. See tähendab, et fumaraat lihtsalt seob enda külge veemolekuli, mille tulemuseks on L-õunhape.

Kaheksas reaktsioon
Krebsi tsükli viimane reaktsioon on L-õunhappe oksüdeerimine oksaloatsetaadiks, st. oksaloäädikhape. Nagu te mõistate, on "oksaloatsetaat" ja "oksaloäädikhape" sünonüümid. Tõenäoliselt mäletate, et oksaloäädikhape on Krebsi tsükli esimese reaktsiooni komponent.
Siinkohal märgime reaktsiooni eripära: NADH 2 moodustumine, mis viib elektronid hingamisahelasse. Ärge unustage ka reaktsioone 3,4 ja 6, seal tekivad ka hingamisahela elektronide ja prootonite kandjad.

Nagu näete, tõstsin punasega konkreetselt esile reaktsioonid, mille käigus moodustuvad NADH ja FADH2. Need on hingamisahela jaoks väga olulised ained. Tõstasin rohelisega esile reaktsiooni, milles toimub substraadi fosforüülimine ja GTP tootmine.
Kuidas seda kõike meeles pidada?
Tegelikult pole see nii raske. Pärast minu kahe artikli, samuti õpiku ja loengute täielikku lugemist peate lihtsalt harjutama nende reaktsioonide kirjutamist. Soovitan meeles pidada Krebsi tsüklit 4 reaktsiooni plokkidena. Kirjutage need 4 reaktsiooni mitu korda, valides igaühe jaoks oma mälule sobiva seose.
Näiteks jäi mulle kohe väga kergesti meelde teine reaktsioon, kus sidrunhappest tekib isotsitrhape (mis on vist kõigile tuttav lapsepõlvest).
Võite kasutada ka mnemoonikaid, näiteks: " Terve ananass ja tükk sufleet on tegelikult minu tänane lõunasöök, mis vastab seeriale - tsitraat, cis-akonitaat, isotsitraat, alfa-ketoglutaraat, suktsinüül-CoA, suktsinaat, fumaraat, malaat, oksaloatsetaat." Selliseid on hunnik veel.
Aga ausalt öeldes ei meeldinud mulle sellised luuletused peaaegu kunagi. Minu arvates on reaktsioonide jada ise lihtsam meeles pidada. Mind aitas palju Krebsi tsükli jagamine kaheks osaks, millest kummaski harjutasin kirjutamist mitu korda tunnis. Reeglina juhtus see sellistes tundides nagu psühholoogia või bioeetika. See on väga mugav – ilma, et teid loengust segataks, saate kulutada sõna otseses mõttes minuti, et kirjutada reaktsioone nii, nagu need teile meenub, ja seejärel kontrollida neid õige valikuga.
Muide, mõnes ülikoolis ei nõua õpetajad biokeemia testide ja eksamite ajal reaktsioonide endi tundmist. Peate lihtsalt teadma, mis on Krebsi tsükkel, kus see toimub, millised on selle omadused ja tähendus ning loomulikult ka transformatsioonide ahel ise. Ainult ahelat saab nimetada ilma valemiteta, kasutades ainult ainete nimetusi. See lähenemine ei ole minu arvates mõttetu.
Loodan, et minu TCA tsükli juhend on teile abiks olnud. Ja ma tahan teile meelde tuletada, et need kaks artiklit ei asenda täielikult teie loenguid ja õpikuid. Kirjutasin need ainult selleks, et saaksite umbkaudu aru, mis Krebsi tsükkel on. Kui näete minu juhendis äkki mõnda viga, kirjutage sellest kommentaarides. Täname tähelepanu eest!
Lühike ajalooline teave
Meie lemmiktsükkel on TCA tsükkel ehk trikarboksüülhappe tsükkel - elu Maal ja Maa all ja Maal... Lõpetage, üldiselt on see kõige hämmastavam mehhanism - see on universaalne, see on viis oksüdeerida süsivesikute, rasvade, valkude laguproduktid elusorganismide rakkudes, mille tulemusena saame energiat oma keha tegevusteks.
Selle protsessi avastas Hans Krebs ise, mille eest ta sai Nobeli preemia!
Ta sündis 25. augustil - 1900 Saksamaa linnas Hildesheimis. Meditsiinihariduse omandas ta Hamburgi ülikoolist ja jätkas biokeemilisi uuringuid Otto Warburgi juhtimisel Berliinis.
1930. aastal avastas ta koos õpilasega organismis ammoniaagi neutraliseerimise protsessi, mis esines paljudel elumaailma esindajatel, sealhulgas inimestel. See tsükkel on uureatsükkel, mida tuntakse ka Krebsi tsüklina nr 1.
Kui Hitler võimule tuli, emigreerus Hans Suurbritanniasse, kus jätkab Cambridge'i ja Sheffieldi ülikoolides loodusteaduste õppimist. Arendades Ungari biokeemiku Albert Szent-Györgyi uurimistööd, sai ta ülevaate ja tegi kuulsaima Krebsi tsükli nr 2 ehk teisisõnu “Szent-Györgyö – Krebsi tsükli” – 1937. aastal.
Uurimistulemused saadetakse ajakirjale Nature, kes keeldub artiklit avaldamast. Seejärel lendab tekst Hollandi ajakirja "Enzymologia". Krebs sai 1953. aastal Nobeli füsioloogia või meditsiini preemia.
Avastus oli üllatav: 1935. aastal leidis Szent-Györgyi, et merevaik-, oksaloäädik-, fumaar- ja õunhape (kõik 4 hapet on loomarakkude looduslikud keemilised komponendid) võimendavad tuvi rinnalihases oksüdatsiooniprotsessi. Mis purustati.
Just selles toimuvad ainevahetusprotsessid suurima kiirusega.
F. Knoop ja K. Martius leidsid 1937. aastal, et sidrunhape muudetakse isotsitrhappeks vaheprodukti, cis - akoniithappe kaudu. Lisaks saab isotsitrhapet muuta a-ketoglutaarhappeks ja selle merevaikhappeks.
Krebs märkas hapete mõju O2 imendumisele tuvi rinnalihases ning tuvastas aktiveeriva toime PVC oksüdatsioonile ja atsetüülkoensüüm A moodustumisele. Lisaks pärssis maloonhape lihastes toimuvaid protsesse. , mis on sarnane merevaikhappega ja võib konkureerivalt inhibeerida ensüüme, mille substraadiks on merevaikhape.
Kui Krebs lisas reaktsioonisegule maloonhapet, algas a-ketoglutaar-, sidrun- ja merevaikhappe kuhjumine. Seega on selge, et a-ketoglutaar- ja sidrunhappe koosmõjul tekib merevaikhape.
Hans uuris rohkem kui 20 muud ainet, kuid need ei mõjutanud oksüdatsiooni. Saadud andmeid võrreldes sai Krebs tsükli. Alguses ei osanud teadlane kindlalt öelda, kas protsess algas sidrun- või isotsitrhappega, mistõttu nimetas ta seda "trikarboksüülhappe tsükliks".
Nüüd teame, et esimene on sidrunhape, nii et õige nimi on tsitraaditsükkel või sidrunhappe tsükkel.
Eukarüootides toimuvad TCA tsükli reaktsioonid mitokondrites, samal ajal kui kõik katalüüsi ensüümid, välja arvatud 1, sisalduvad mitokondriaalses maatriksis vabas olekus; erand on suktsinaatdehüdrogenaas, mis paikneb mitokondrite sisemembraanil ja on sisseehitatud lipiidide kaksikkiht. Prokarüootides toimuvad tsükli reaktsioonid tsütoplasmas.
Kohtume tsüklis osalejatega:
1) Atsetüülkoensüüm A:- atsetüülrühm
- koensüüm A - koensüüm A:
2) PIKE – oksaloatsetaat – oksaloäädikhape:
näib koosnevat kahest osast: oksaal- ja äädikhappest.
3-4) Sidrun- ja isotsitrhape:
5) a-ketoglutaarhape:
6) suktsinüül-koensüüm A:
7) Merevaikhape:
8) fumaarhape:
9) õunhape:
Kuidas reaktsioonid tekivad? Üldiselt oleme me kõik harjunud sõrmuse välimusega, mis on näha alloleval pildil. Allpool kirjeldatakse kõike samm-sammult:
1. Atsetüülkoensüümi A ja oksaloäädikhappe ➙ sidrunhappe kondensatsioon.
Atsetüülkoensüümi A muundumine algab kondenseerumisega oksaloäädikhappega, mille tulemusena moodustub sidrunhape.
Reaktsioon ei nõua ATP tarbimist, kuna energia selle protsessi jaoks saadakse tioeetri sideme hüdrolüüsi tulemusena atsetüülkoensüümiga A, mis on kõrge energiaga:
2. Sidrunhape läheb läbi cis-akoniithappe isotsitrhappeks.
Toimub sidrunhappe isomerisatsioon isotsitrhappeks. Konversiooniensüüm - akonitaas - dehüdreerib esmalt sidrunhapet, moodustades cis-akoniithappe, seejärel ühendab vee metaboliidi kaksiksidemega, moodustades isotsitrhappe:
3. Isotsitrhapet dehüdrogeenitakse, moodustades α-ketoglutaarhapet ja CO2.
Isotsitrhapet oksüdeerib spetsiifiline dehüdrogenaas, mille koensüümiks on NAD.
Samaaegselt oksüdatsiooniga toimub isotsitrhappe dekarboksüülimine. Transformatsioonide tulemusena tekib α-ketoglutaarhape.
4. Alfa-ketoglutaarhapet dehüdrogeenivad ➙ suktsinüülkoensüüm A ja CO2.
Järgmine etapp on α-ketoglutaarhappe oksüdatiivne dekarboksüülimine.
Katalüüsib α-ketoglutaraadi dehüdrogenaasi kompleks, mis on mehhanismi, struktuuri ja toime poolest sarnane püruvaadi dehüdrogenaasi kompleksiga. Selle tulemusena moodustub suktsinüül-CoA.
5. Suktsinüülkoensüüm A ➙ merevaikhape.
Suktsinüül-CoA hüdrolüüsitakse vabaks merevaikhappeks, vabanev energia salvestatakse guanosiintrifosfaadi moodustumisel. See etapp on tsüklis ainuke, kus energiat otseselt vabaneb.
6. Merevaikhape on dehüdrogeenitud ➙ fumaarhape.
Merevaikhappe dehüdrogeenimist kiirendab suktsinaatdehüdrogenaas, selle koensüümiks on FAD.
7. Fumaarhape on hüdraatunud ➙ õunhape.
Merevaikhappe dehüdrogeenimisel tekkiv fumaarhape hüdraatub ja moodustub õunhape.
8. Õunhape on dehüdrogeenitud ➙ Oksaal-äädikhape – tsükkel sulgub.
Lõplik protsess on õunhappe dehüdrogeenimine, mida katalüüsib malaatdehüdrogenaas;
Etapi tulemuseks on metaboliit, millega trikarboksüülhappe tsükkel algab – oksaal-äädikhape.
Järgmise tsükli reaktsioonis 1 siseneb veel üks kogus atsetüülkoensüümi A.
Kuidas seda tsüklit meeles pidada? Lihtsalt!
1) Väga kujundlik väljend:Terve ananass ja tükk sufleet on tegelikult minu tänane lõunasöök, mis vastab - tsitraat, cis-akonitaat, isotsitraat, (alfa-)ketoglutaraat, suktsinüül-CoA, suktsinaat, fumaraat, malaat, oksaloatsetaat.
2) Veel üks pikk luuletus:
PIKE sõi atsetaati, selgus, et tsitraat,
Tsisakonitaadi kaudu muutub see isotsitraadiks.
Pärast vesiniku loovutamist NAD-ile kaotab see CO2,
Alfa-ketoglutaraat on selle üle ülimalt õnnelik.
Oksüdatsioon on tulemas – NAD varastas vesiniku,
TDP, koensüüm A võtab CO2.
Ja vaevu ilmus suktsinüülis energia,
Kohe sündis ATP ja allesjäänud oli suktsinaat.
Nüüd jõudis ta FAD-i - ta vajab vesinikku,
Fumaraat jõi veest ja muutus malaadiks.
Siis jõudis NAD malaadini, omandas vesiniku,
HAUGI ilmus uuesti ja peitis end vaikselt.
3) Algne luuletus – lühidalt:
HAUGI ATSETÜLLIMONIL,
Kuid hobune kartis nartsissi,
Ta on temast üle ISOLIMON
ALFA – KETOGLUTARASED.
KOENSÜÜMIGA SUKKINALISEERITUD,
AMBER FUMAROVO,
Varusin talveks mõned õunad,
Muutus jälle HAUGIKS.
TRIKARBOKSÜÜLHAPETE TÜKKL (KREBS TÜKKEL)
Glükolüüs muudab glükoosi püruvaadiks ja toodab glükoosimolekulist kaks ATP molekuli – väikese osa selle molekuli potentsiaalsest energiast.
Aeroobsetes tingimustes muudetakse püruvaat glükolüüsist atsetüül-CoA-ks ja oksüdeeritakse trikarboksüülhappe tsüklis (sidrunhappe tsükkel) CO2-ks. Sel juhul läbivad selle tsükli reaktsioonides vabanenud elektronid NADH ja FADH 2 kuni 0 2 - lõpliku aktseptorini. Elektronide transport on seotud prootoni gradiendi tekkega mitokondriaalses membraanis, mille energiat kasutatakse seejärel oksüdatiivse fosforüülimise tulemusena ATP sünteesiks. Vaatleme neid reaktsioone.
Aeroobsetes tingimustes läbib püroviinamarihape (1. etapp) oksüdatiivse dekarboksüülimise, mis on tõhusam kui muundamine piimhappeks, mille käigus moodustub atsetüül-CoA (2. etapp), mida saab oksüdeerida glükoosi lagunemise lõppsaadusteks - CO 2 ja H 2 0 (3. etapp). Saksa biokeemik G. Krebs (1900-1981), uurinud üksikute orgaaniliste hapete oksüdatsiooni, ühendas nende reaktsioonid üheks tsükliks. Seetõttu nimetatakse trikarboksüülhappe tsüklit tema auks sageli Krebsi tsükliks.
Püruviinhappe oksüdatsioon atsetüül-CoA-ks toimub mitokondrites kolme ensüümi (püruvaatdehüdrogenaas, lipoamiiddehüdrogenaas, lipoüülatsetüültransferaas) ja viie koensüümi (NAD, FAD, tiamiinpürofosfaat, lipoehappeamiid, koensüüm A) osalusel. Need neli koensüümi sisaldavad B-vitamiine (B x, B 2, B 3, B 5), mis viitab nende vitamiinide vajadusele süsivesikute normaalseks oksüdatsiooniks. Selle keerulise ensüümsüsteemi mõjul muundatakse püruvaat oksüdatiivse dekarboksüülimisreaktsiooni käigus äädikhappe aktiivseks vormiks - atsetüülkoensüümiks A:
Füsioloogilistes tingimustes on püruvaatdehüdrogenaas eranditult pöördumatu ensüüm, mis seletab rasvhapete süsivesikuteks muutmise võimatust.
Kõrge energiaga sideme olemasolu atsetüül-CoA molekulis näitab selle ühendi suurt reaktsioonivõimet. Eelkõige võib atsetüül-CoA toimida mitokondrites energia tootmiseks; maksas kasutatakse liigset atsetüül-CoA-d ketokehade sünteesiks; tsütosoolis osaleb see keeruliste molekulide, nagu steroidid ja rasvhapped, sünteesis.
Püroviinamarihappe oksüdatiivse dekarboksüülimise reaktsioonis saadud atsetüül-CoA siseneb trikarboksüülhappe tsüklisse (Krebsi tsükkel). Krebsi tsükkel, süsivesikute, rasvade ja aminohapete oksüdatsiooni viimane kataboolne rada, on sisuliselt "ainevahetuse pada". Krebsi tsükli reaktsioone, mis toimuvad eranditult mitokondrites, nimetatakse ka sidrunhappe tsükliks või trikarboksüülhappe tsükliks (TCA tsükkel).
Trikarboksüülhappe tsükli üks olulisemaid funktsioone on redutseeritud koensüümide genereerimine (3 molekuli NADH + H + ja 1 molekul FADH 2), millele järgneb vesinikuaatomite või nende elektronide ülekandmine lõplikule aktseptorile - molekulaarsele hapnikule. Selle transpordiga kaasneb vaba energia suur vähenemine, millest osa kasutatakse oksüdatiivse fosforüülimise protsessis ATP kujul säilitamiseks. On selge, et trikarboksüülhappe tsükkel on aeroobne, hapnikust sõltuv.
1. Trikarboksüülhappe tsükli esialgne reaktsioon on atsetüül-CoA ja oksaloäädikhappe kondenseerumine mitokondriaalse maatriksi ensüümi tsitraadi süntaasi osalusel, moodustades sidrunhappe.

2. Tsitraadist veemolekuli eemaldamist katalüüsiva ensüümi akonitaas mõjul pöördub viimane

cis-akoonhappeks. Vesi ühineb cis-akoniithappega, muutudes isotsitrhappeks.
3. Ensüüm isotsitraatdehüdrogenaas katalüüsib seejärel sidrunhappe tsükli esimest dehüdrogenaasi reaktsiooni, kui isotsitrhape muudetakse oksüdatiivse dekarboksüülimise teel α-ketoglutaarhappeks:

Selles reaktsioonis moodustub esimene CO 2 molekul ja NADH 4- H + tsükli esimene molekul.
4. α-ketoglutaarhappe edasist muundamist suktsinüül-CoA-ks katalüüsib α-ketoglutaardehüdrogenaasi multiensüümne kompleks. See reaktsioon on keemiliselt analoogne püruvaadi dehüdrogenaasi reaktsiooniga. See hõlmab lipoehapet, tiamiinpürofosfaati, HS-KoA, NAD +, FAD. 
Selle reaktsiooni tulemusena moodustub taas NADH + H + ja CO 2 molekul.
5. Suktsinüül-CoA molekulil on kõrge energiaga side, mille energia salvestub järgmises reaktsioonis GTP kujul. Ensüüm suktsinüül-CoA süntetaasi mõjul muundatakse suktsinüül-CoA vabaks merevaikhappeks. Pange tähele, et merevaikhapet võib saada ka metüülmalonüül-CoA-st paaritu arvu süsinikuaatomitega rasvhapete oksüdeerimisel.

See reaktsioon on näide substraadi fosforüülimisest, kuna suure energiaga GTP molekul moodustub sel juhul ilma elektronide ja hapniku transpordiahela osaluseta.
6. Merevaikhape oksüdeeritakse suktsinaatdehüdrogenaasi reaktsioonis fumaarhappeks. Suktsinaatdehüdrogenaas, tüüpiline rauda väävlit sisaldav ensüüm, mille koensüümiks on FAD. Suktsinaatdehüdrogenaas on ainus ensüüm, mis on ankurdatud sisemise mitokondriaalse membraani külge, samas kui kõik teised tsükli ensüümid asuvad mitokondriaalses maatriksis. 
7. Sellele järgneb fumaarhappe hüdratiseerimine õunhappeks ensüümi fumaraas toimel füsioloogilistes tingimustes pöörduvas reaktsioonis:

8. Trikarboksüülhappe tsükli lõppreaktsioon on aktiivse ensüümi mitokondriaalse NAD~-sõltuva malaatdehüdrogenaasi osalusel malaatdehüdrogenaasi reaktsioon, milles moodustub kolmas redutseeritud NADH + H + molekul:

Oksaloäädikhappe (oksaloatsetaadi) moodustumine lõpetab trikarboksüülhappe tsükli ühe pöörde. Oksalaäädikhapet saab kasutada atsetüül-CoA teise molekuli oksüdeerimiseks ja seda reaktsioonitsüklit saab korrata mitu korda, mis viib pidevalt oksaloäädikhappe tootmiseni.
Seega põhjustab ühe atsetüül-CoA molekuli oksüdeerimine TCA tsüklis tsükli substraadina ühe GTP molekuli, kolme NADP + H + molekuli ja ühe FADH 2 molekuli tootmise. Nende redutseerivate ainete oksüdeerimine bioloogilises oksüdatsiooniahelas

leensioon viib 12 ATP molekuli sünteesini. See arvutus tuleb selgelt välja teemast “Bioloogiline oksüdatsioon”: ühe NAD + molekuli kaasamisega elektronide transpordisüsteemi kaasneb lõpuks 3 ATP molekuli moodustumine, FADH 2 molekuli kaasamine tagab 2 ATP molekuli moodustumise, ja üks GTP molekul võrdub 1 ATP molekuliga.
Pange tähele, et kaks adetüül-CoA süsinikuaatomit sisenevad trikarboksüülhappe tsüklisse ja kaks süsinikuaatomit lahkuvad tsüklist CO2-na dekarboksüülimisreaktsioonides, mida katalüüsivad isotsitraatdehüdrogenaas ja alfa-ketoglutaraatdehüdrogenaas.
Glükoosi molekuli täielikul oksüdeerumisel aeroobsetes tingimustes C0 2 ja H 2 0-ks tekib energia ATP kujul:
- 4 ATP molekuli glükoosimolekuli muundamisel 2 püroviinamarihappe molekuliks (glükolüüs);
- 6 ATP molekuli, mis tekkisid 3-fosfoglütseraldehüüddehüdrogenaasi reaktsioonis (glükolüüs);
- 30 ATP molekuli, mis tekkisid kahe püroviinamarihappe molekuli oksüdeerumisel püruvaadi dehüdrogenaasi reaktsioonis ja sellele järgnenud kahe atsetüül-CoA molekuli muundumisel CO 2 -ks ja H 2 0 -ks trikarboksüülhappe tsüklis. Seetõttu võib glükoosimolekuli täieliku oksüdeerumise koguenergia olla 40 ATP molekuli. Siiski tuleb arvestada, et glükoosi oksüdatsiooni käigus kulub kaks ATP molekuli glükoosi glükoos-6-fosfaadiks ja fruktoos-6-fosfaadi fruktoos-1,6-ks muutmise etapis. difosfaat. Seetõttu on glükoosimolekuli oksüdatsioonist saadav "neto" energia 38 ATP molekuli.
Saate võrrelda glükoosi anaeroobse glükolüüsi ja aeroobse katabolismi energeetikat. 1 grammis glükoosi molekulis (180 g) teoreetiliselt sisalduvast 688 kcal energiast on 20 kcal kahes anaeroobse glükolüüsi reaktsioonides tekkinud ATP molekulis ja 628 kcal jääb teoreetiliselt piimhappe kujule.
Aeroobsetes tingimustes saadakse 688 kcal grammi glükoosi molekulist 38 ATP molekulis 380 kcal. Seega on glükoosi kasutamise efektiivsus aeroobsetes tingimustes ligikaudu 19 korda kõrgem kui anaeroobsel glükolüüsil.
Tuleb märkida, et kõik oksüdatsioonireaktsioonid (trioosfosfaadi oksüdatsioon, püroviinamarihape, neli trikarboksüülhappe tsükli oksüdatsioonireaktsiooni) konkureerivad ATP sünteesil ADP-st ja fosforist (Pasteuri efekt). See tähendab, et oksüdatsioonireaktsioonides tekkival molekulil NADH + H + on valida hingamissüsteemi reaktsioonide vahel, mis kannavad vesinikku hapnikuks, ja ensüümi LDH vahel, mis kannab vesinikku üle püroviinamarihappele.
Trikarboksüülhappe tsükli varases staadiumis võivad selle happed tsüklist lahkuda, et osaleda teiste rakuühendite sünteesis ilma tsükli enda toimimist häirimata. Trikarboksüülhappe tsükli aktiivsuse reguleerimisega on seotud erinevad tegurid. Nende hulgas tuleks mainida eelkõige atsetüül-CoA molekulide varu, püruvaatdehüdrogenaasi kompleksi aktiivsust, hingamisahela komponentide aktiivsust ja sellega seotud oksüdatiivset fosforüülimist, aga ka oksaloäädikhappe taset.
Molekulaarne hapnik ei osale otseselt trikarboksüülhappe tsüklis, kuid selle reaktsioonid toimuvad ainult aeroobsetes tingimustes, kuna NAD ~ ja FAD saab mitokondrites regenereerida ainult elektronide ülekandmisel molekulaarsesse hapnikku. Tuleb rõhutada, et erinevalt trikarboksüülhappe tsüklist on glükolüüs võimalik ka anaeroobsetes tingimustes, kuna NAD~ regenereeritakse püroviinamarihappe üleminekul piimhappeks.
Lisaks ATP moodustumisele on trikarboksüülhappe tsüklil veel üks oluline tähendus: tsükkel annab vahestruktuure erinevatele organismi biosünteesidele. Näiteks enamik porfüriinide aatomeid pärineb suktsinüül-CoA-st, paljud aminohapped on α-ketoglutaar- ja oksaloäädikhappe derivaadid ning fumaarhape esineb uurea sünteesi protsessis. See näitab trikarboksüülhappe tsükli terviklikkust süsivesikute, rasvade ja valkude metabolismis.
Nagu näitavad glükolüüsi reaktsioonid, peitub enamiku rakkude võime energiat genereerida nende mitokondrites. Mitokondrite arv erinevates kudedes on seotud kudede füsioloogiliste funktsioonidega ja peegeldab nende võimet osaleda aeroobsetes tingimustes. Näiteks punalibledel puuduvad mitokondrid ja seetõttu ei ole neil võimet genereerida energiat, kasutades lõpliku elektroni aktseptorina hapnikku. Aeroobsetes tingimustes töötava südamelihase puhul on aga pool raku tsütoplasma mahust esindatud mitokondrite poolt. Maks sõltub oma erinevate funktsioonide täitmiseks ka aeroobsetest tingimustest ja imetajate hepatotsüüdid sisaldavad kuni 2 tuhat mitokondrit raku kohta.
Mitokondrid sisaldavad kahte membraani - välimist ja sisemist. Välismembraan on lihtsam, koosneb 50% rasvadest ja 50% valkudest ning sellel on suhteliselt vähe funktsioone. Sisemembraan on struktuurselt ja funktsionaalselt keerulisem. Ligikaudu 80% selle mahust moodustavad valgud. See sisaldab enamikku ensüümidest, mis osalevad elektronide transpordis ja oksüdatiivses fosforüülimises, metaboolseid vaheühendeid ja adeniini nukleotiide tsütosooli ja mitokondriaalse maatriksi vahel.
Erinevad redoksreaktsioonides osalevad nukleotiidid, nagu NAD +, NADH, NADP +, FAD ja FADH 2, ei tungi läbi sisemise mitokondriaalse membraani. Atsetüül-CoA ei saa liikuda mitokondriaalsest sektsioonist tsütosooli, kus see on vajalik rasvhapete või steroolide sünteesiks. Seetõttu muundatakse intramitokondriaalne atsetüül-CoA trikarboksüülhappe tsükli tsitraadi süntaasi reaktsiooniks ja siseneb sellisel kujul tsütosooli.
Trikarboksüülhappe tsükli avastas esmakordselt inglise biokeemik Krebs. Ta oli esimene, kes postuleeris selle tsükli tähtsuse püruvaadi täielikuks põlemiseks, mille peamiseks allikaks on süsivesikute glükolüütiline muundamine. Seejärel näidati, et trikarboksüülhappe tsükkel on "fookus", kus peaaegu kõik metaboolsed rajad lähenevad.
Niisiis siseneb püruvaadi oksüdatiivse dekarboksüülimise tulemusena moodustunud atsetüül-CoA Krebsi tsüklisse. See tsükkel koosneb kaheksast järjestikusest reaktsioonist (joonis 91). Tsükkel algab atsetüül-CoA kondenseerumisega oksaloatsetaadiga ja sidrunhappe moodustumisega. ( Nagu allpool näha, ei oksüdeeru tsüklis mitte atsetüül-CoA ise, vaid keerulisem ühend - sidrunhape (trikarboksüülhape).)
Seejärel kaotab sidrunhape (kuue süsiniku ühend) läbi mitme dehüdrogeenimise (vesiniku eemaldamine) ja dekarboksüülimise (CO 2 eemaldamine) kaks süsinikuaatomit ja jälle ilmub Krebsi tsüklisse oksaloatsetaat (nelja süsiniku ühend). st tsükli täieliku pöörde tulemusena põleb atsetüül-CoA molekul CO 2 ja H 2 O-ks ning oksaloatsetaadi molekul regenereeritakse. Allpool on kõik kaheksa Krebsi tsükli järjestikust reaktsiooni (etappi).
Esimeses reaktsioonis, mida katalüüsib ensüüm tsitraadi süntaas, kondenseeritakse atsetüül-CoA oksaloatsetaadiga. Selle tulemusena moodustub sidrunhape:

Ilmselt moodustub selles reaktsioonis vaheproduktina ensüümiga seotud tsitriil-CoA. Viimane hüdrolüüsitakse seejärel spontaanselt ja pöördumatult, moodustades tsitraadi ja HS-CoA.
Tsükli teises reaktsioonis läbib saadud sidrunhape dehüdratsiooni, moodustades cis-akoniithappe, mis veemolekuli lisamisel muutub isotsitrhappeks. Neid pöörduvaid hüdratatsiooni-dehüdratatsiooni reaktsioone katalüüsib ensüüm akonitaathüdraas:

Kolmandas reaktsioonis, mis näib olevat Krebsi tsükli kiirust piirav reaktsioon, dehüdrogeenitakse isotsitrhape NAD-sõltuva isotsitraatdehüdrogenaasi juuresolekul:

(Kudedes on kahte tüüpi isotsitraatdehüdrogenaase: NAD- ja NADP-sõltuvad. On kindlaks tehtud, et NAD-sõltuv isotsitraatdehüdrogenaas mängib Krebsi tsüklis isotsitrhappe oksüdatsiooni peamise katalüsaatori rolli.)
Isotsitraadi dehüdrogenaasi reaktsiooni käigus isotsitrhape dekarboksüleeritakse. NAD-sõltuv isotsitraatdehüdrogenaas on allosteeriline ensüüm, mis vajab spetsiifilise aktivaatorina ADP-d. Lisaks vajab ensüüm oma aktiivsuse avaldamiseks Mg 2+ või Mn 2+ ioone.
Neljandas reaktsioonis dekarboksüülitakse a-ketoglutaarhape oksüdatiivselt suktsinüül-CoA-ks. Selle reaktsiooni mehhanism on sarnane püruvaadi oksüdatiivse dekarboksüülimise reaktsiooniga atsetüül-CoA-ks. α-ketoglutaraadi dehüdrogenaasi kompleks on struktuurilt sarnane püruvaadi dehüdrogenaasi kompleksiga. Mõlemal juhul osaleb reaktsioonis viis koensüümi: TDP, lipoehappe amiid, HS-CoA, FAD ja NAD. Kokkuvõttes saab selle reaktsiooni kirjutada järgmiselt:

Viiendat reaktsiooni katalüüsib ensüüm suktsinüül-CoA süntetaas. Selle reaktsiooni käigus muudetakse suktsinüül-CoA GDP ja anorgaanilise fosfaadi osalusel merevaikhappeks (suktsinaadiks). Samal ajal tekib suktsinüül-CoA kõrge energiasisaldusega tioestersideme tõttu GTP1 kõrge energiasisaldusega fosfaatside:

(Saadud GTP annetab seejärel oma terminaalse fosfaatrühma ADP-le, mille tulemusena moodustub ATP. Kõrge energiaga nukleosiidtrifosfaadi moodustumine suktsinüül-CoA süntetaasi reaktsiooni käigus on näide fosforüülimisest substraadi tasemel.)
Kuuendas reaktsioonis dehüdrogeenistatakse suktsinaat fumaarhappeks. Suktsinaadi oksüdatsiooni katalüüsib suktsinaatdehüdrogenaas, mille molekulis on koensüüm FAD kovalentselt seotud valguga:

Seitsmendas reaktsioonis hüdraaditakse saadud fumaarhape ensüümi fumaraathüdrataasi toimel. Selle reaktsiooni saadus on õunhape (malaat). Tuleb märkida, et fumaraathüdrataas on stereospetsiifiline - selle reaktsiooni käigus moodustub L-õunhape:

Lõpuks oksüdeeritakse trikarboksüülhappe tsükli kaheksandas reaktsioonis mitokondriaalse NAD-sõltuva malaadi dehüdrogenaasi mõjul L-malaat oksaloatsetaadiks:

Nagu näete, toimub kaheksast ensümaatilisest reaktsioonist koosneva tsükli ühes pöördes ühe atsetüül-CoA molekuli täielik oksüdatsioon (“põlemine”). Tsükli pidevaks toimimiseks on vajalik pidev atsetüül-CoA varustamine süsteemi ning redutseeritud olekusse läinud koensüümid (NAD ja FAD) tuleb ikka ja jälle oksüdeerida. See oksüdatsioon toimub mitokondrites asuvas elektronide transpordisüsteemis (või hingamisteede ensüümide ahelas).
Atsetüül-CoA oksüdatsiooni tulemusena vabanev energia on suures osas kontsentreeritud ATP kõrge energiasisaldusega fosfaatsidemetesse. Neljast vesinikuaatomipaarist kolm paari kantakse NAD kaudu elektronide transpordisüsteemi; sel juhul moodustub bioloogilise oksüdatsioonisüsteemi iga paari kohta kolm ATP molekuli (konjugeeritud oksüdatiivse fosforüülimise protsessis) ja seega kokku üheksa ATP molekuli. Üks aatomipaar siseneb FAD-i kaudu elektronide transpordisüsteemi, mille tulemusena moodustub 2 ATP molekuli. Krebsi tsükli reaktsioonide käigus sünteesitakse ka 1 molekul GTP-d, mis võrdub 1 molekuliga ATP-ga. Seega tekitab atsetüül-CoA oksüdatsioon Krebsi tsüklis 12 ATP molekuli.

Nagu juba märgitud, moodustub püruvaadi oksüdatiivsel dekarboksüülimisel atsetüül-CoA-ks 1 molekul NADH2 (3 molekuli ATP-d). Kuna ühe glükoosimolekuli lagunemisel tekib kaks püruvaadi molekuli, siis kui need oksüdeeritakse kaheks atsetüül-CoA molekuliks ja järgneval kahel trikarboksüülhappe tsüklil, sünteesitakse 30 ATP molekuli (seega ühe molekuli oksüdatsioon). püruvaadi muutmine CO 2 -ks ja H 2 O-ks tekitab 15 ATP molekuli).
Sellele tuleb lisada 2 aeroobse glükolüüsi käigus tekkinud ATP molekuli ja 4 ekstramitokondriaalse NADH 2 molekuli oksüdatsioonil sünteesitud ATP molekuli, mis tekivad 2 molekuli glütseraldehüüd-3-fosfaadi oksüdatsiooni käigus dehüdrogenaasi reaktsioonis. Kokku leiame, et kui 1 glükoosimolekul laguneb kudedes vastavalt võrrandile: C 6 H 12 0 6 + 60 2 -> 6CO 2 + 6H 2 O, sünteesitakse 36 ATP molekuli, mis aitab kaasa glükoosi akumuleerumisele. adenosiintrifosfaat suure energiaga fosfaatsidemetes 36 X 34,5 ~ 1240 kJ (või muude allikate järgi 36 X 38 ~ 1430 kJ) vaba energiat. Ehk siis kogu glükoosi aeroobsel oksüdatsioonil vabanevast vabast energiast (umbes 2840 kJ) koguneb kuni 50% sellest mitokondritesse kujul, mida saab kasutada erinevate füsioloogiliste funktsioonide täitmiseks. Pole kahtlust, et energeetiliselt on glükoosi täielik lagundamine tõhusam protsess kui glükolüüs. Tuleb märkida, et glütseraldehüüd-3-fosfaat 2 muundamisel moodustunud NADH 2 molekulid ei tekita järgneval oksüdatsioonil mitte 6 ATP molekuli, vaid ainult 4. Fakt on see, et ekstramitokondriaalse NADH 2 molekulid ise ei suuda tungivad läbi membraani mitokondritesse. Nende annetatud elektronid saab aga kaasata bioloogilise oksüdatsiooni mitokondriaalsesse ahelasse, kasutades niinimetatud glütserofosfaadi süstikumehhanismi (joonis 92). Nagu jooniselt näha, reageerib tsütoplasmaatiline NADH2 esmalt tsütoplasmaatilise dihüdroksüatsetoonfosfaadiga, moodustades glütserool-3-fosfaadi. Reaktsiooni katalüüsib NAD-sõltuv tsütoplasmaatiline glütserool-3-fosfaatdehüdrogenaas.